
Carnosine, Small but Mighty—Prospect of Use as Functional Ingredient for Functional Food Formulation

 ,
, 
 , , ,
, , ,  , ,
, ,  , , ,
, , ,  and add
Show full author list
and add
Show full author list
Abstract
:1. Introduction
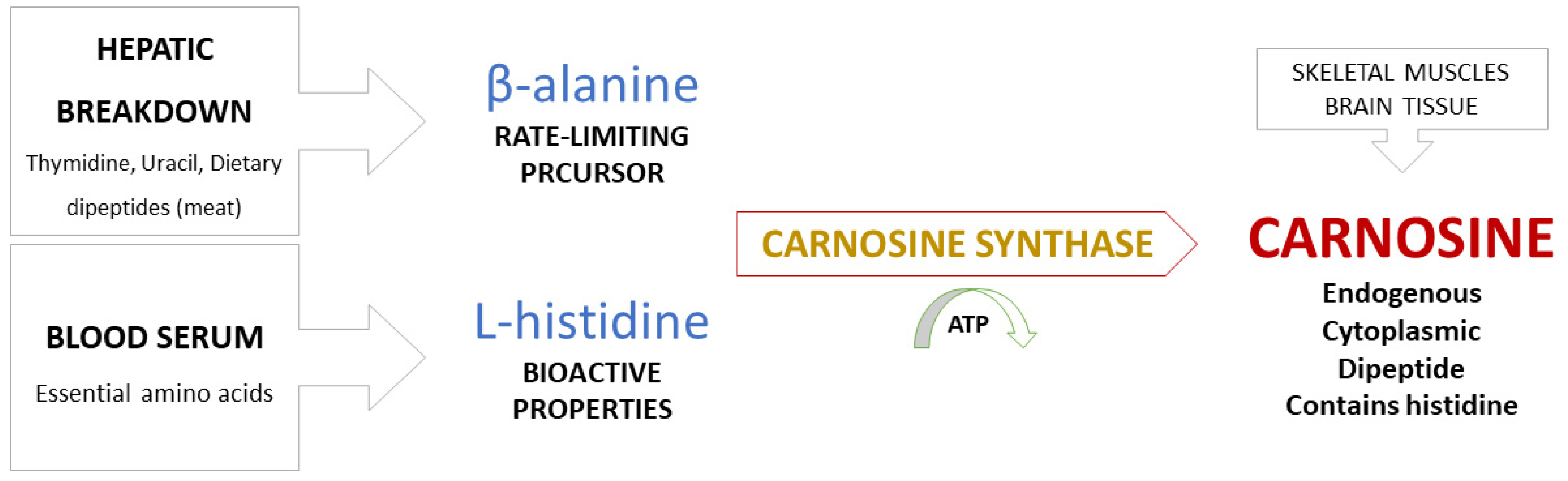
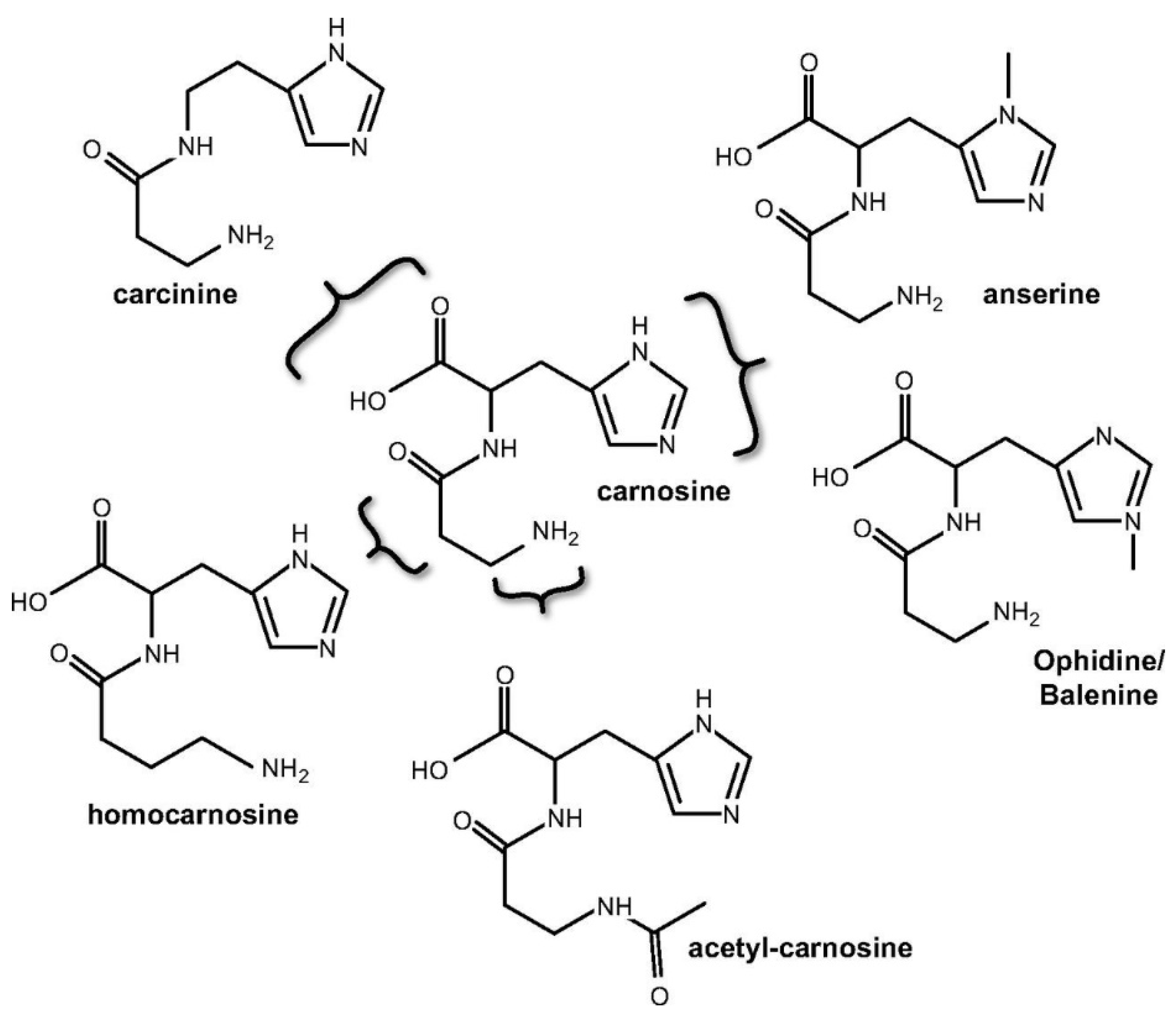
2. Carnosine Metabolism
3. Methodology of Carnosine Determination
Analytical Methods for Carnosine Determination
4. Challenges in Carnosine Examination
5. Carnosine Biological Effects
5.1. Carnosine in Muscle Function
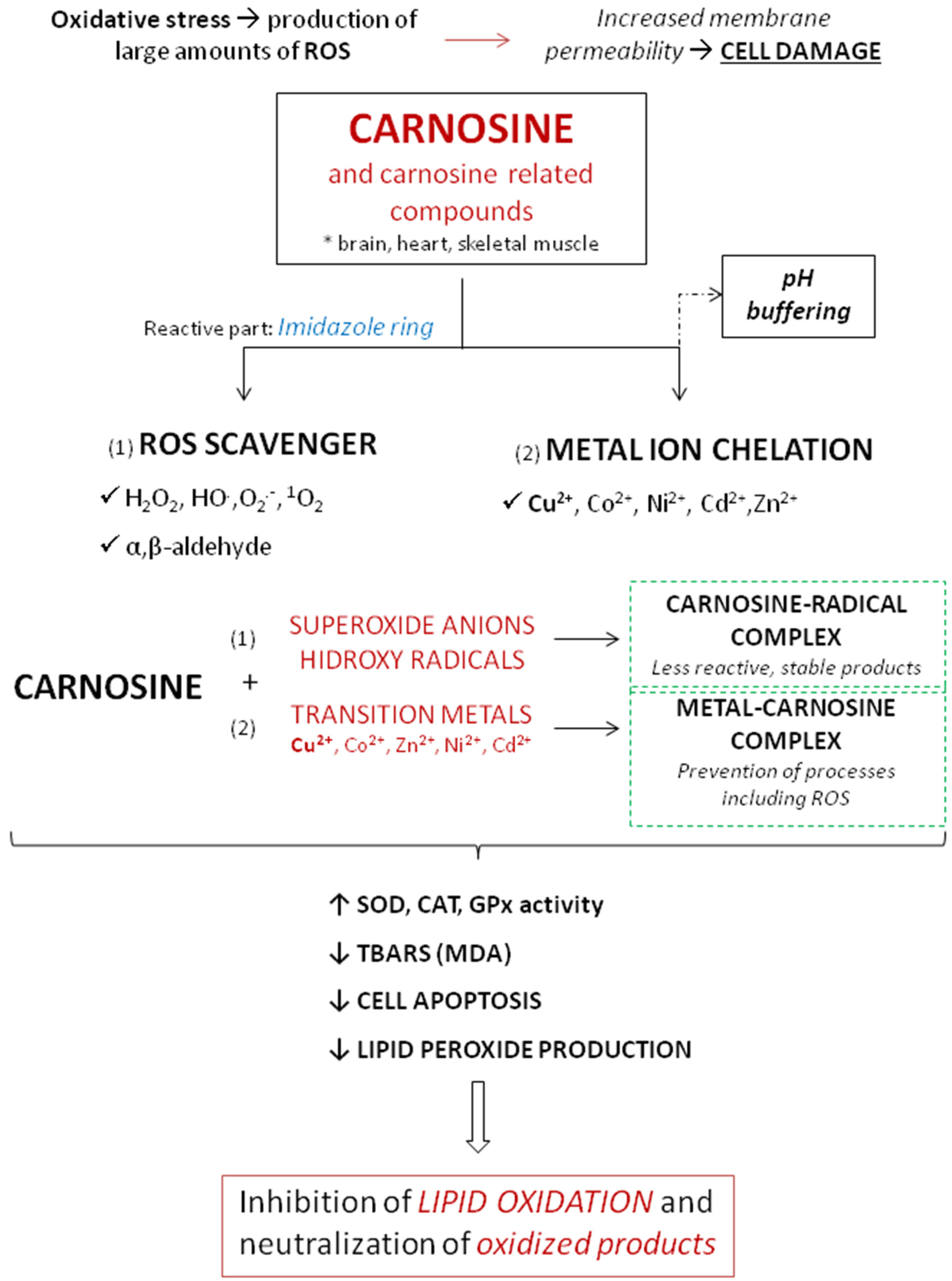
5.2. Antioxidant and Anti-Inflammatory Properties of Carnosine
5.3. Immunomodulation
5.4. Carnosine and Aging
6. Carnosine Supplementation
7. Forthcoming Research—Potential Health Benefits and Clinical Use
7.1. Carnosine in Glucose Metabolism/Diabetes
7.2. Carnosine in Cardiovascular Disorders (Atherosclerosis, Heart Failure)
7.3. Carnosine in Neurological Disorders
7.4. Carnosine in Malignant Diseases
7.5. Carnosine and Vascular Function
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gulewitsch, W.; Amiradžibi, S. Ueber das Carnosin, eine neue organische Base des Fleischextractes. Berichte der Dtsch. Chem. Gesellschaft 1900, 33, 1902–1903. [Google Scholar] [CrossRef] [Green Version]
- Aristoy, M.C.; Toldrá, F. Histidine dipeptides HPLC-based test for the detection of mammalian origin proteins in feeds for ruminants. Meat Sci. 2004, 67, 211–217. [Google Scholar] [CrossRef]
- Boldyrev, A.A.; Aldini, G.; Derave, W. Physiology and Pathophysiology of Carnosine. Physiol. Rev. 2013, 93, 1803–1845. [Google Scholar] [CrossRef]
- Chasovnikova, L.V.; Formazyuk, V.E.; Sergienko, V.I.; Boldyrev, A.A.; Severin, S.E. The antioxidative properties of carnosine and other drugs. Biochem. Int. 1990, 20, 1097–1103. [Google Scholar]
- Bellia, F.; Vecchio, G.; Cuzzocrea, S.; Calabrese, V.; Rizzarelli, E. Neuroprotective features of carnosine in oxidative driven diseases. Mol. Aspects Med. 2011, 32, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Chee, M.E.; Zhang, H.; Zhang, W.; Mine, Y. Carnosine—a natural bioactive dipeptide: Bioaccessibility, bioavailability and health benefits. J. Food Bioact. 2019, 5, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Vistoli, G.; Straniero, V.; Pedretti, A.; Fumagalli, L.; Bolchi, C.; Pallavicini, M.; Valoti, E.; Testa, B. Predicting the physicochemical profile of diastereoisomeric histidine-containing dipeptides by property space analysis. Chirality 2012, 24, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Torreggiani, A.; Tamba, M.; Fini, G. Binding of copper(II) to carnosine: Raman and IR spectroscopic study. Biopolymers 2000, 57, 149–159. [Google Scholar] [CrossRef]
- Hipkiss, A.R. Carnosine and Its Possible Roles in Nutrition and Health. Adv. Food Nutr. Res. 2009, 57, 87–154. [Google Scholar] [PubMed]
- Ghodsi, R.; Kheirouri, S. Carnosine and advanced glycation end products: A systematic review. Amino Acids 2018, 50, 1177–1186. [Google Scholar] [CrossRef]
- Kim, M.Y.; Kim, E.J.; Kim, Y.-N.; Choi, C.; Lee, B.-H. Effects of α-lipoic acid and L-carnosine supplementation on antioxidant activities and lipid profiles in rats. Nutr. Res. Pract. 2011, 5, 421. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.Y.; Jiang, Z.Y.; Lin, Y.C.; Zheng, C.T.; Zhou, G.L. Dietary supplementation with carnosine improves antioxidant capacity and meat quality of finishing pigs. J. Anim. Physiol. Anim. Nutr. 2010, 94, e286–e295. [Google Scholar] [CrossRef]
- Derave, W.; Everaert, I.; Beeckman, S.; Baguet, A. Muscle Carnosine Metabolism and β-Alanine Supplementation in Relation to Exercise and Training. Sport. Med. 2010, 40, 247–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decker, E.A.; Faraji, H. Inhibition of lipid oxidation by carnosine. J. Am. Oil Chem. Soc. 1990, 67, 650–652. [Google Scholar] [CrossRef]
- Ferraris, R.P.; Diamond, J.; Kwan, W.W. Dietary regulation of intestinal transport of the dipeptide carnosine. Am. J. Physiol. Liver Physiol. 1988, 255, G143–G150. [Google Scholar] [CrossRef]
- Gardner, M.L.; Illingworth, K.M.; Kelleher, J.; Wood, D. Intestinal absorption of the intact peptide carnosine in man, and comparison with intestinal permeability to lactulose. J. Physiol. 1991, 439, 411–422. [Google Scholar] [CrossRef]
- Fernández-García, E.; Carvajal-Lérida, I.; Pérez-Gálvez, A. In vitro bioaccessibility assessment as a prediction tool of nutritional efficiency. Nutr. Res. 2009, 29, 751–760. [Google Scholar] [CrossRef]
- Park, Y.J.; Volpe, S.L.; Decker, E.A. Quantitation of Carnosine in Humans Plasma after Dietary Consumption of Beef. J. Agric. Food Chem. 2005, 53, 4736–4739. [Google Scholar] [CrossRef]
- Marcolini, E.; Babini, E.; Bordoni, A.; Di Nunzio, M.; Laghi, L.; Maczó, A.; Picone, G.; Szerdahelyi, E.; Valli, V.; Capozzi, F. Bioaccessibility of the Bioactive Peptide Carnosine during in Vitro Digestion of Cured Beef Meat. J. Agric. Food Chem. 2015, 63, 4973–4978. [Google Scholar] [CrossRef] [PubMed]
- Ansurudeen, I.; Sunkari, V.G.; Grünler, J.; Peters, V.; Schmitt, C.P.; Catrina, S.-B.; Brismar, K.; Forsberg, E.A. Carnosine enhances diabetic wound healing in the db/db mouse model of type 2 diabetes. Amino Acids 2012, 43, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Boldyrev, A.A.; Stvolinsky, S.L.; Fedorova, T.N.; Suslina, Z.A. Carnosine As a Natural Antioxidant and Geroprotector: From Molecular Mechanisms to Clinical Trials. Rejuvenation Res. 2010, 13, 156–158. [Google Scholar] [CrossRef]
- Maherani, B.; Arab-Tehrany, E.; Kheirolomoom, A.; Cleymand, F.; Linder, M. Influence of lipid composition on physicochemical properties of nanoliposomes encapsulating natural dipeptide antioxidant l-carnosine. Food Chem. 2012, 134, 632–640. [Google Scholar] [CrossRef]
- Bellia, F.; Vecchio, G.; Rizzarelli, E. Carnosine derivatives: New multifunctional drug-like molecules. Amino Acids 2012, 43, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Bellia, F.; Vecchio, G.; Rizzarelli, E. Carnosinases, Their Substrates and Diseases. Molecules 2014, 19, 2299–2329. [Google Scholar] [CrossRef] [Green Version]
- Baye, E.; Ukropcova, B.; Ukropec, J.; Hipkiss, A.; Aldini, G.; de Courten, B. Physiological and therapeutic effects of carnosine on cardiometabolic risk and disease. Amino Acids 2016, 48, 1131–1149. [Google Scholar] [CrossRef] [PubMed]
- Pavlin, M.; Rossetti, G.; De Vivo, M.; Carloni, P. Carnosine and Homocarnosine Degradation Mechanisms by the Human Carnosinase Enzyme CN1: Insights from Multiscale Simulations. Biochemistry 2016, 55, 2772–2784. [Google Scholar] [CrossRef]
- Peters, V.; Klessens, C.Q.F.; Baelde, H.J.; Singler, B.; Veraar, K.A.M.; Zutinic, A.; Drozak, J.; Zschocke, J.; Schmitt, C.P.; de Heer, E. Intrinsic carnosine metabolism in the human kidney. Amino Acids 2015, 47, 2541–2550. [Google Scholar] [CrossRef] [Green Version]
- Peters, V.; Jansen, E.E.W.; Jakobs, C.; Riedl, E.; Janssen, B.; Yard, B.A.; Wedel, J.; Hoffmann, G.F.; Zschocke, J.; Gotthardt, D.; et al. Anserine inhibits carnosine degradation but in human serum carnosinase (CN1) is not correlated with histidine dipeptide concentration. Clin. Chim. Acta 2011, 412, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Schön, M.; Mousa, A.; Berk, M.; Chia, W.L.; Ukropec, J.; Majid, A.; Ukropcová, B.; de Courten, B. The Potential of Carnosine in Brain-Related Disorders: A Comprehensive Review of Current Evidence. Nutrients 2019, 11, 1196. [Google Scholar] [CrossRef] [Green Version]
- Budzeń, S.; Rymaszewska, J. The biological role of carnosine and its possible applications in medicine. Adv. Clin. Exp. Med. 2013, 22, 739–744. [Google Scholar]
- Gariballa, S. Review. Carnosine: Physiological properties and therapeutic potential. Age Ageing 2000, 29, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Harris, R.C.; Dunnett, M.; Greenhaff, P.L. Carnosine and taurine contents in individual fibres of human vastus lateralis muscle. J. Sports Sci. 1998, 16, 639–643. [Google Scholar] [CrossRef]
- Dunnett, M.; Harris, R.C. High-performance liquid chromatographic determination of imidazole dipeptides, histidine, 1-methylhistidine and 3-methylhistidine in equine and camel muscle and individual muscle fibres. J. Chromatogr. B Biomed. Sci. Appl. 1997, 688, 47–55. [Google Scholar] [CrossRef]
- Hill, C.A.; Harris, R.C.; Kim, H.J.; Harris, B.D.; Sale, C.; Boobis, L.H.; Kim, C.K.; Wise, J.A. Influence of β-alanine supplementation on skeletal muscle carnosine concentrations and high intensity cycling capacity. Amino Acids 2007, 32, 225–233. [Google Scholar] [CrossRef]
- Kendrick, I.P.; Kim, H.J.; Harris, R.C.; Kim, C.K.; Dang, V.H.; Lam, T.Q.; Bui, T.T.; Wise, J.A. The effect of 4 weeks β-alanine supplementation and isokinetic training on carnosine concentrations in type I and II human skeletal muscle fibres. Eur. J. Appl. Physiol. 2009, 106, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Ito, O.; Mukai, N.; Takahashi, H.; Takamatsu, K. High Level of Skeletal Muscle Carnosine Contributes to the Latter Half of Exercise Performance during 30-s Maximal Cycle Ergometer Sprinting. Jpn. J. Physiol. 2002, 52, 199–205. [Google Scholar] [CrossRef] [Green Version]
- BLANCQUAERT, L.; EVERAERT, I.; MISSINNE, M.; BAGUET, A.; STEGEN, S.; VOLKAERT, A.; PETROVIC, M.; VERVAET, C.; ACHTEN, E.; DE MAEYER, M.; et al. Effects of Histidine and β-alanine Supplementation on Human Muscle Carnosine Storage. Med. Sci. Sport. Exerc. 2017, 49, 602–609. [Google Scholar] [CrossRef]
- Keytsman, C.; Blancquaert, L.; Wens, I.; Missine, M.; Van Noten, P.; Vandenabeele, F.; Derave, W.; Eijnde, B.O. Muscle carnosine in experimental autoimmune encephalomyelitis and multiple sclerosis. Mult. Scler. Relat. Disord. 2018, 21, 24–29. [Google Scholar] [CrossRef] [Green Version]
- Varanoske, A.N.; Hoffman, J.R.; Church, D.D.; Coker, N.A.; Baker, K.M.; Dodd, S.J.; Harris, R.C.; Oliveira, L.P.; Dawson, V.L.; Wang, R.; et al. Comparison of sustained-release and rapid-release β-alanine formulations on changes in skeletal muscle carnosine and histidine content and isometric performance following a muscle-damaging protocol. Amino Acids 2019, 51, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Pandya, V.K.; Sonwane, B.; Rathore, R.; Unnikrishnan, A.G.; Kumaran, S.; Kulkarni, M.J. Development of multiple reaction monitoring assay for quantification of carnosine in human plasma. RSC Adv. 2020, 10, 763–769. [Google Scholar] [CrossRef] [Green Version]
- Everaert, I.; Taes, Y.; De Heer, E.; Baelde, H.; Zutinic, A.; Yard, B.; Sauerhöfer, S.; Vanhee, L.; Delanghe, J.; Aldini, G.; et al. Low plasma carnosinase activity promotes carnosinemia after carnosine ingestion in humans. Am. J. Physiol. Physiol. 2012, 302, F1537–F1544. [Google Scholar] [CrossRef] [PubMed]
- Manyam, B.V. Cerebrospinal Fluid Amino Compounds in Parkinson’s Disease. Arch. Neurol. 1988, 45, 48. [Google Scholar] [CrossRef] [PubMed]
- Fontes, J.E.d.N.; Rodriguez-Caturla, M.Y.; Sant’Ana, A.S.; Lopes, T.I.B.; Marsaioli, A.J. Foodomics and storage monitoring of three meat cuts by 1 H NMR. Concepts Magn. Reson. Part A 2018, 47A, e21474. [Google Scholar] [CrossRef] [Green Version]
- Solis, M.Y.; Cooper, S.; Hobson, R.M.; Artioli, G.G.; Otaduy, M.C.; Roschel, H.; Robertson, J.; Martin, D.; Painelli, V.S.; Harris, R.C.; et al. Effects of Beta-Alanine Supplementation on Brain Homocarnosine/Carnosine Signal and Cognitive Function: An Exploratory Study. PLoS ONE 2015, 10, e0123857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özdemir, M.S.; Reyngoudt, H.; De Deene, Y.; Sazak, H.S.; Fieremans, E.; Delputte, S.; D’Asseler, Y.; Derave, W.; Lemahieu, I.; Achten, E. Absolute quantification of carnosine in human calf muscle by proton magnetic resonance spectroscopy. Phys. Med. Biol. 2007, 52, 6781–6794. [Google Scholar] [CrossRef]
- Noblitt, S.D.; Staicu, L.C.; Ackerson, C.J.; Henry, C.S. Sensitive, Selective Analysis of Selenium Oxoanions Using Microchip Electrophoresis with Contact Conductivity Detection. Anal. Chem. 2014, 86, 8425–8432. [Google Scholar] [CrossRef] [Green Version]
- Fekete, S.; Guillarme, D.; Sandra, P.; Sandra, K. Chromatographic, Electrophoretic, and Mass Spectrometric Methods for the Analytical Characterization of Protein Biopharmaceuticals. Anal. Chem. 2016, 88, 480–507. [Google Scholar] [CrossRef]
- HUANG, Y.; DUAN, J.; ZHANG, J.; CHEN, G. Online Sample Stacking for Determination of Carnosine-Related Peptides by Capillary Electrophoresis. Chinese J. Chromatogr. 2007, 25, 326–332. [Google Scholar] [CrossRef]
- Staňová, A.; Marák, J.; Rezeli, M.; Páger, C.; Kilár, F.; Kaniansky, D. Analysis of therapeutic peptides in human urine by combination of capillary zone electrophoresis–electrospray mass spectrometry with preparative capillary isotachophoresis sample pretreatment. J. Chromatogr. A 2011, 1218, 8701–8707. [Google Scholar] [CrossRef]
- Zhao, S.; Huang, Y.; Shi, M.; Huang, J.; Liu, Y.-M. Quantification of carnosine-related peptides by microchip electrophoresis with chemiluminescence detection. Anal. Biochem. 2009, 393, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Jozanović, M.; Hajduković, M.; Galović, O.; Kralik, G.; Kralik, Z.; Sakač, N.; Medvidović-Kosanović, M.; Sak-Bosnar, M. Determination of anti-oxidative histidine dipeptides in poultry by microchip capillary electrophoresis with contactless conductivity detection. Food Chem. 2017, 221, 1658–1665. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Shi, Y.; Duan, J.; Chen, G. Field-amplified on-line sample stacking for determination of carnosine-related peptides by capillary electrophoresis. J. Sep. Sci. 2006, 29, 1026–1030. [Google Scholar] [CrossRef] [PubMed]
- Ragab, M.A.A.; El-Kimary, E.I. Recent Advances and Applications of Microfluidic Capillary Electrophoresis: A Comprehensive Review (2017–Mid 2019). Crit. Rev. Anal. Chem. 2020, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Lara, F.J.; Airado-Rodríguez, D.; Moreno-González, D.; Huertas-Pérez, J.F.; García-Campaña, A.M. Applications of capillary electrophoresis with chemiluminescence detection in clinical, environmental and food analysis. A review. Anal. Chim. Acta 2016, 913, 22–40. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.E.; Kaleta, E.J.; Arora, A.; Wolk, D.M. Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry: A Fundamental Shift in the Routine Practice of Clinical Microbiology. Clin. Microbiol. Rev. 2013, 26, 547–603. [Google Scholar] [CrossRef] [Green Version]
- Pisarev, D.; Novikov, O.; Vasiliev, G.; Selyutin, O. Experience in using the MALDI/TOF/MS method in pharmaceutical analysis. Sci. Rep. Belgorod State Univ. Ser. Med. Pharmacy 2012, 18, 1–11. [Google Scholar]
- Uenoyama, R.; Miyazaki, M.; Miyazaki, T.; Shigeno, Y.; Tokairin, Y.; Konno, H.; Yamashita, T. LC-ESI-MS/MS quantification of carnosine, anserine, and balenine in meat samples. J. Chromatogr. B 2019, 1132, 121826. [Google Scholar] [CrossRef]
- Jozanović, M.; Medvidović-Kosanović, M.; Sak-Bosnar, M. Voltammetric Characterization and Determination of Histidine Dipeptides-Carnosine and Anserine. Int. J. Electrochem. Sci. 2015, 10, 6548–6557. [Google Scholar]
- Wu, G. Important roles of dietary taurine, creatine, carnosine, anserine and 4-hydroxyproline in human nutrition and health. Amino Acids 2020, 52, 329–360. [Google Scholar] [CrossRef] [Green Version]
- Wu, G. Dietary protein intake and human health. Food Funct. 2016, 7, 1251–1265. [Google Scholar] [CrossRef] [Green Version]
- Szterk, A.; Roszko, M. Simultaneous determination of free amino acids, l-carnosine, purine, pyrimidine, and nucleosides in meat by liquid chromatography/single quadrupole mass spectrometry. J. Liq. Chromatogr. Relat. Technol. 2014, 37, 664–680. [Google Scholar] [CrossRef]
- Clifford, W.M. The Effect of Cold Storage on the Carnosine Content of Muscle. Biochem. J. 1922, 16, 341–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sale, C.; Saunders, B.; Harris, R.C. Effect of beta-alanine supplementation on muscle carnosine concentrations and exercise performance. Amino Acids 2010, 39, 321–333. [Google Scholar] [CrossRef]
- Harris, R.C.; Wise, J.A.; Price, K.A.; Kim, H.J.; Kim, C.K.; Sale, C. Determinants of muscle carnosine content. Amino Acids 2012, 43, 5–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MATTHEWS, J.J.; ARTIOLI, G.G.; TURNER, M.D.; SALE, C. The Physiological Roles of Carnosine and β-Alanine in Exercising Human Skeletal Muscle. Med. Sci. Sport. Exerc. 2019, 51, 2098–2108. [Google Scholar] [CrossRef] [PubMed]
- GOODALL, M.C. Carnosine Phosphate as Phosphate Donor in Muscular Contraction. Nature 1956, 178, 539–540. [Google Scholar] [CrossRef]
- Severin, S.E.; Kirzon, M.V.; Kaftanova, T.M. Effect of carnosine and anserine on action of isolated frog muscles. Dokl. Akad. Nauk SSSR 1953, 91, 691–694. [Google Scholar]
- Boldyrev, A.A. Carnosine: New concept for the function of an old molecule. Biochem. 2012, 77, 313–326. [Google Scholar] [CrossRef]
- Smith, E.C.B. The buffering of muscle in rigor; protein, phosphate and carnosine. J. Physiol. 1938, 92, 336–343. [Google Scholar] [CrossRef] [Green Version]
- Deutsch, A.; Eggleton, P. The titration constants of anserine, carnosine and some related compounds. Biochem. J. 1938, 32, 209–211. [Google Scholar] [CrossRef]
- Saunders, B.; Elliott-Sale, K.; Artioli, G.G.; Swinton, P.A.; Dolan, E.; Roschel, H.; Sale, C.; Gualano, B. β-alanine supplementation to improve exercise capacity and performance: A systematic review and meta-analysis. Br. J. Sports Med. 2017, 51, 658–669. [Google Scholar] [CrossRef]
- Culbertson, J.Y.; Kreider, R.B.; Greenwood, M.; Cooke, M. Effects of Beta-Alanine on Muscle Carnosine and Exercise Performance: A Review of the Current Literature. Nutrients 2010, 2, 75–98. [Google Scholar] [CrossRef] [Green Version]
- Junge, W.; McLaughlin, S. The role of fixed and mobile buffers in the kinetics of proton movement. Biochim. Biophys. Acta Bioenerg. 1987, 890, 1–5. [Google Scholar] [CrossRef]
- Swietach, P.; Youm, J.-B.; Saegusa, N.; Leem, C.-H.; Spitzer, K.W.; Vaughan-Jones, R.D. Coupled Ca2+/H+ transport by cytoplasmic buffers regulates local Ca2+ and H+ ion signaling. Proc. Natl. Acad. Sci. USA 2013, 110, E2064–E2073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baguet, A.; Koppo, K.; Pottier, A.; Derave, W. β-Alanine supplementation reduces acidosis but not oxygen uptake response during high-intensity cycling exercise. Eur. J. Appl. Physiol. 2010, 108, 495–503. [Google Scholar] [CrossRef]
- Dawson, R., Jr.; Biasetti, M.; Messina, S.; Dominy, J. The cytoprotective role of taurine in exercise-induced muscle injury. Amino Acids 2002, 22, 309–324. [Google Scholar] [CrossRef]
- Boldyrev, A.A.; Dupin, A.M.; Pindel, E.V.; Severin, S.E. Antioxidative properties of histidine-containing dipeptides from skeletal muscles of vertebrates. Comp. Biochem. Physiol. Part B Comp. Biochem. 1988, 89, 245–250. [Google Scholar] [CrossRef]
- Gordon, A.M.; Homsher, E.; Regnier, M. Regulation of Contraction in Striated Muscle. Physiol. Rev. 2000, 80, 853–924. [Google Scholar] [CrossRef]
- Batrukova, M.A.; Rubtsov, A.M. Histidine-containing dipeptides as endogenous regulators of the activity of sarcoplasmic reticulum Ca-release channels. Biochim. Biophys. Acta Biomembr. 1997, 1324, 142–150. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Song, L.; Cheng, X.; Yang, Y.; Luan, B.; Jia, L.; Xu, F.; Zhang, Z. Carnosine pretreatment protects against hypoxia–ischemia brain damage in the neonatal rat model. Eur. J. Pharmacol. 2011, 667, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Lamb, G.D.; Stephenson, D.G. Effect of Mg2+ on the control of Ca2+ release in skeletal muscle fibres of the toad. J. Physiol. 1991, 434, 507–528. [Google Scholar] [CrossRef]
- Dutka, T.L.; Lamb, G.D. Effect of Carnosine on Excitation–Contraction Coupling in Mechanically-Skinned Rat Skeletal Muscle. J. Muscle Res. Cell Motil. 2004, 25, 203–213. [Google Scholar] [CrossRef]
- Dutka, T.L.; Lamboley, C.R.; McKenna, M.J.; Murphy, R.M.; Lamb, G.D. Effects of carnosine on contractile apparatus Ca 2+ sensitivity and sarcoplasmic reticulum Ca 2+ release in human skeletal muscle fibers. J. Appl. Physiol. 2012, 112, 728–736. [Google Scholar] [CrossRef] [Green Version]
- Lamont, C.; Miller, D.J. Calcium sensitizing action of carnosine and other endogenous imidazoles in chemically skinned striated muscle. J. Physiol. 1992, 454, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Bolitho Donaldson, S.K.; Hermansen, L.; Bolles, L. Differential, direct effects of H+ on Ca2+-activated force of Skinned fibers from the soleus, cardiac and adductor magnus muscles of rabbits. Pflügers Arch. Eur. J. Physiol. 1978, 376, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.L.; Barnett, C.T.; Davidson, J.; Maritza, B.; Fraser, W.D.; Harris, R.; Sale, C. β-alanine supplementation improves in-vivo fresh and fatigued skeletal muscle relaxation speed. Eur. J. Appl. Physiol. 2017, 117, 867–879. [Google Scholar] [CrossRef] [Green Version]
- Tsugorka, A.; Rios, E.; Blatter, L. Imaging elementary events of calcium release in skeletal muscle cells. Science 1995, 269, 1723–1726. [Google Scholar] [CrossRef] [PubMed]
- Tallon, M.J.; Harris, R.C.; Boobis, L.H.; Fallowfield, J.L.; Wise, J.A. The Carnosine Content of Vastus Lateralis Is Elevated in Resistance-Trained Bodybuilders. J. Strength Cond. Res. 2005, 19, 725. [Google Scholar] [CrossRef] [PubMed]
- Nagai, K.; Niijima, A.; Yamano, T.; Otani, H.; Okumra, N.; Tsuruoka, N.; Nakai, M.; Kiso, Y. Possible Role of L-Carnosine in the Regulation of Blood Glucose through Controlling Autonomic Nerves. Exp. Biol. Med. 2003, 228, 1138–1145. [Google Scholar] [CrossRef]
- Nordsborg, N.; Mohr, M.; Pedersen, L.D.; Nielsen, J.J.; Langberg, H.; Bangsbo, J. Muscle interstitial potassium kinetics during intense exhaustive exercise: Effect of previous arm exercise. Am. J. Physiol. Integr. Comp. Physiol. 2003, 285, R143–R148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greene, S.M.; Margolis, F.L.; Grillo, M.; Fisher, H. Enhanced carnosine (β-alanyl-L-histidine) breakdown and histamine metabolism following treatment with compound. Eur. J. Pharmacol. 1984, 99, 79–84. [Google Scholar] [CrossRef]
- IKEDA, T.; KIMURA, K.; HAMA, T.; TAMAKI, N. Activation of Rabbit Muscle Fructose 1,6-Bisphosphatase by Histidine and Carnosine. J. Biochem. 1980, 87, 179–185. [Google Scholar] [CrossRef]
- Brown, C.E. Interactions among carnosine, anserine, ophidine and copper in biochemical adaptation. J. Theor. Biol. 1981, 88, 245–256. [Google Scholar] [CrossRef]
- de Andrade Kratz, C.; de Salles Painelli, V.; de Andrade Nemezio, K.M.; da Silva, R.P.; Franchini, E.; Zagatto, A.M.; Gualano, B.; Artioli, G.G. Beta-alanine supplementation enhances judo-related performance in highly-trained athletes. J. Sci. Med. Sport 2017, 20, 403–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobias, G.; Benatti, F.B.; de Salles Painelli, V.; Roschel, H.; Gualano, B.; Sale, C.; Harris, R.C.; Lancha, A.H.; Artioli, G.G. Additive effects of beta-alanine and sodium bicarbonate on upper-body intermittent performance. Amino Acids 2013, 45, 309–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, R.P.; de Oliveira, L.F.; Saunders, B.; de Andrade Kratz, C.; de Salles Painelli, V.; da Eira Silva, V.; Marins, J.C.B.; Franchini, E.; Gualano, B.; Artioli, G.G. Effects of β-alanine and sodium bicarbonate supplementation on the estimated energy system contribution during high-intensity intermittent exercise. Amino Acids 2019, 51, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Jordan, T.; Lukaszuk, J.; Misic, M.; Umoren, J. Effect of beta-alanine supplementation on the onset of blood lactate accumulation (OBLA) during treadmill running: Pre/post 2 treatment experimental design. J. Int. Soc. Sports Nutr. 2010, 7, 20. [Google Scholar] [CrossRef] [Green Version]
- Everaert, I.; Mooyaart, A.; Baguet, A.; Zutinic, A.; Baelde, H.; Achten, E.; Taes, Y.; De Heer, E.; Derave, W. Vegetarianism, female gender and increasing age, but not CNDP1 genotype, are associated with reduced muscle carnosine levels in humans. Amino Acids 2011, 40, 1221–1229. [Google Scholar] [CrossRef]
- Asatoor, A.M.; Bandoh, J.K.; Lant, A.F.; Milne, M.D.; Navab, F. Intestinal absorption of carnosine and its constituent amino acids in man. Gut 1970, 11, 250–254. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Xu, J.; Cui, D.; Liu, L.; Zhang, S.; Shen, B.; Wu, Y.; Zhang, Q. Protective effect of carnosine on hydrogen peroxide–induced oxidative stress in human kidney tubular epithelial cells. Biochem. Biophys. Res. Commun. 2021, 534, 576–582. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 2015; ISBN 9780198717478. [Google Scholar]
- Pavlov, A.R.; Revina, A.A.; Dupin, A.M.; Boldyrev, A.A.; Yaropolov, A.I. The mechanism of interaction of carnosine with superoxide radicals in water solutions. Biochim. Biophys. Acta Gen. Subj. 1993, 1157, 304–312. [Google Scholar] [CrossRef]
- Torreggiani, M.T.A. A pulse radiolysis study of carnosine in aqueous solution. Int. J. Radiat. Biol. 1998, 74, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Miller, D. Transition metals as catalysts of “autoxidation” reactions. Free Radic. Biol. Med. 1990, 8, 95–108. [Google Scholar] [CrossRef]
- Hartman, P.E.; Hartman, Z.; Ault, K.T. Scavenging of singlet molecular oxygen by imidazole compounds: High and sustained activities of carboxy terminal histidine dipeptides and exceptional activity of imidazole-4-acetic acid. Photochem. Photobiol. 1990, 51, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Baran, E.J. Metal complexes of carnosine. Biochemistry 2000, 65, 789–797. [Google Scholar] [PubMed]
- Torreggiani, M.; Tamba, A. Hydroxyl radical scavenging by carnosine and Cu(II)-carnosine complexes: A pulse-radiolysis and spectroscopic study. Int. J. Radiat. Biol. 1999, 75, 1177–1188. [Google Scholar] [CrossRef] [PubMed]
- Crush, K. Carnosine and related substances in animal tissues. Comp. Biochem. Physiol. 1970, 34, 3–30. [Google Scholar] [CrossRef]
- Decker, E.A.; Livisay, S.A.; Zhou, S. A re-evaluation of the antioxidant activity of purified carnosine. Biochemistry 2000, 65, 766–770. [Google Scholar]
- Aldini, G.; Orioli, M.; Rossoni, G.; Savi, F.; Braidotti, P.; Vistoli, G.; Yeum, K.-J.; Negrisoli, G.; Carini, M. The carbonyl scavenger carnosine ameliorates dyslipidaemia and renal function in Zucker obese rats. J. Cell. Mol. Med. 2011, 15, 1339–1354. [Google Scholar] [CrossRef]
- Klebanov, G.I.; Teselkin, Y.O.; Babenkova, I.V.; Popov, I.N.; Levin, G.; Tyulina, O.V.; Boldyrev, A.A.; Vladimirov, Y.A. Evidence for a direct interaction of superoxide anion radical with carnosine. IUBMB Life 1997, 43, 99–106. [Google Scholar] [CrossRef]
- Aydın, A.F.; Küçükgergin, C.; Özdemirler-Erata, G.; Koçak-Toker, N.; Uysal, M. The effect of carnosine treatment on prooxidant–antioxidant balance in liver, heart and brain tissues of male aged rats. Biogerontology 2010, 11, 103–109. [Google Scholar] [CrossRef]
- Aldini, G.; Facino, R.M.; Beretta, G.; Carini, M. Carnosine and related dipeptides as quenchers of reactive carbonyl species: From structural studies to therapeutic perspectives. BioFactors 2005, 24, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef]
- Carletto, C.; Nicolay, J.F.; Courbebaisse, Y. Courbebaisse Oxidative stress and cutaneous ageing: The ‘toxic second messengers’ concept and an interesting family of products, ‘pseudodipeptides. ’ Int. J. Cosmet. Sci. 2000, 22, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Poli, G.; Schaur, R.J.; Siems, W.G.; Leonarduzzi, G. 4-Hydroxynonenal: A membrane lipid oxidation product of medicinal interest. Med. Res. Rev. 2008, 28, 569–631. [Google Scholar] [CrossRef]
- Shoeb, M.; Ansari, N.; Srivastava, S.; Ramana, K. 4-Hydroxynonenal in the Pathogenesis and Progression of Human Diseases. Curr. Med. Chem. 2013, 21, 230–237. [Google Scholar] [CrossRef]
- Schaur, R.; Siems, W.; Bresgen, N.; Eckl, P. 4-Hydroxy-nonenal—A Bioactive Lipid Peroxidation Product. Biomolecules 2015, 5, 2247–2337. [Google Scholar] [CrossRef] [Green Version]
- Barski, O.A.; Xie, Z.; Baba, S.P.; Sithu, S.D.; Agarwal, A.; Cai, J.; Bhatnagar, A.; Srivastava, S. Dietary Carnosine Prevents Early Atherosclerotic Lesion Formation in Apolipoprotein E–Null Mice. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1162–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frohnert, B.I.; Sinaiko, A.R.; Serrot, F.J.; Foncea, R.E.; Moran, A.; Ikramuddin, S.; Choudry, U.; Bernlohr, D.A. Increased Adipose Protein Carbonylation in Human Obesity. Obesity 2011, 19, 1735–1741. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S.; Yamada, S.; Kashima, M.; Ihara, Y.; Yamada, Y.; Tanaka, T.; Hiai, H.; Seino, Y.; Uchida, K. Serum 4-Hydroxy-2-Nonenal-Modified Albumin Is Elevated in Patients with Type 2 Diabetes Mellitus. Antioxid. Redox Signal. 2000, 2, 681–685. [Google Scholar] [CrossRef]
- Katunga, L.A.; Gudimella, P.; Efird, J.T.; Abernathy, S.; Mattox, T.A.; Beatty, C.; Darden, T.M.; Thayne, K.A.; Alwair, H.; Kypson, A.P.; et al. Obesity in a model of gpx4 haploinsufficiency uncovers a causal role for lipid-derived aldehydes in human metabolic disease and cardiomyopathy. Mol. Metab. 2015, 4, 493–506. [Google Scholar] [CrossRef]
- Anderson, E.J.; Vistoli, G.; Katunga, L.A.; Funai, K.; Regazzoni, L.; Monroe, T.B.; Gilardoni, E.; Cannizzaro, L.; Colzani, M.; De Maddis, D.; et al. A carnosine analog mitigates metabolic disorders of obesity by reducing carbonyl stress. J. Clin. Invest. 2018, 128, 5280–5293. [Google Scholar] [CrossRef]
- Caruso, G.; Fresta, C.; Musso, N.; Giambirtone, M.; Grasso, M.; Spampinato, S.; Merlo, S.; Drago, F.; Lazzarino, G.; Sortino, M.; et al. Carnosine Prevents Aβ-Induced Oxidative Stress and Inflammation in Microglial Cells: A Key Role of TGF-β1. Cells 2019, 8, 64. [Google Scholar] [CrossRef] [Green Version]
- Onufriev, M.V.; Potanova, G.I.; Silaeva, S.A.; Nikolaev, A.I. Carnosine as a stimulator of cytotoxic and phagocytic function of peritoneal macrophages. Biokhimiia 1992, 57, 1352–1359. [Google Scholar] [PubMed]
- Tanaka, K.-I.; Sugizaki, T.; Kanda, Y.; Tamura, F.; Niino, T.; Kawahara, M. Preventive Effects of Carnosine on Lipopolysaccharide-induced Lung Injury. Sci. Rep. 2017, 7, 42813. [Google Scholar] [CrossRef]
- Tsai, S.-J.; Kuo, W.-W.; Liu, W.-H.; Yin, M.-C. Antioxidative and Anti-Inflammatory Protection from Carnosine in the Striatum of MPTP-Treated Mice. J. Agric. Food Chem. 2010, 58, 11510–11516. [Google Scholar] [CrossRef] [PubMed]
- Invernizzi, P.L.; Venerando, B.; Di Pierro, F.; Saronni, S.; Papini, N. Effects of Carnosine and Beta-Alanine Ingestion on Anaerobic Sprint Performance and Peripheral Blood Mononuclear Cell Interleukin-6 and -10 Gene Expression. Adv. Phys. Educ. 2013, 03, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Houjeghani, S.; Kheirouri, S.; Faraji, E.; Jafarabadi, M.A. L-Carnosine supplementation attenuated fasting glucose, triglycerides, advanced glycation end products, and tumor necrosis factor– α levels in patients with type 2 diabetes: A double-blind placebo-controlled randomized clinical trial. Nutr. Res. 2018, 49, 96–106. [Google Scholar] [CrossRef]
- Katakura, Y.; Totsuka, M.; Imabayashi, E.; Matsuda, H.; Hisatsune, T. Anserine/Carnosine Supplementation Suppresses the Expression of the Inflammatory Chemokine CCL24 in Peripheral Blood Mononuclear Cells from Elderly People. Nutrients 2017, 9, 1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasanein, P.; Felegari, Z. Chelating effects of carnosine in ameliorating nickel-induced nephrotoxicity in rats. Can. J. Physiol. Pharmacol. 2017, 95, 1426–1432. [Google Scholar] [CrossRef] [PubMed]
- Navarro, J.F.; Mora, C. Diabetes, Inflammation, Proinflammatory Cytokines, and Diabetic Nephropathy. Sci. World J. 2006, 6, 908–917. [Google Scholar] [CrossRef]
- Stentz, F.B.; Umpierrez, G.E.; Cuervo, R.; Kitabchi, A.E. Proinflammatory Cytokines, Markers of Cardiovascular Risks, Oxidative Stress, and Lipid Peroxidation in Patients With Hyperglycemic Crises. Diabetes 2004, 53, 2079–2086. [Google Scholar] [CrossRef] [Green Version]
- Guest, C.B.; Park, M.J.; Johnson, D.R.; Freund, G.G. The implication of proinflammatory cytokines in type 2 diabetes. Front. Biosci. 2008, Volume, 5187. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Hsu, C.; Lin, M.; Liu, K.; Yin, M. Histidine and carnosine delay diabetic deterioration in mice and protect human low density lipoprotein against oxidation and glycation. Eur. J. Pharmacol. 2005, 513, 145–150. [Google Scholar] [CrossRef] [PubMed]
- de Courten, B.; Jakubova, M.; de Courten, M.P.; Kukurova, I.J.; Vallova, S.; Krumpolec, P.; Valkovic, L.; Kurdiova, T.; Garzon, D.; Barbaresi, S.; et al. Effects of carnosine supplementation on glucose metabolism: Pilot clinical trial. Obesity 2016, 24, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Baye, E.; Ukropec, J.; de Courten, M.P.; Vallova, S.; Krumpolec, P.; Kurdiova, T.; Aldini, G.; Ukropcova, B.; de Courten, B. Effect of carnosine supplementation on the plasma lipidome in overweight and obese adults: A pilot randomised controlled trial. Sci. Rep. 2017, 7, 17458. [Google Scholar] [CrossRef]
- Baye, E.; Ukropec, J.; de Courten, M.P.J.; Kurdiova, T.; Krumpolec, P.; Fernández-Real, J.-M.; Aldini, G.; Ukropcova, B.; de Courten, B. Carnosine supplementation reduces plasma soluble transferrin receptor in healthy overweight or obese individuals: A pilot randomised trial. Amino Acids 2019, 51, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Katsanos, K.H.; Papadakis, K.A. Inflammatory Bowel Disease: Updates on Molecular Targets for Biologics. Gut Liver 2017, 11, 455–463. [Google Scholar] [CrossRef] [Green Version]
- Son, D.O.; Satsu, H.; Kiso, Y.; Shimizu, M. Characterization of carnosine uptake and its physiological function in human intestinal epithelial Caco-2 cells. BioFactors 2004, 21, 395–398. [Google Scholar] [CrossRef]
- Son, D.O.; Satsu, H.; Kiso, Y.; Totsuka, M.; Shimizu, M. Inhibitory effect of carnosine on interleukin-8 production in intestinal epithelial cells through translational regulation. Cytokine 2008, 42, 265–276. [Google Scholar] [CrossRef]
- Hisamatsu, T.; Ono, N.; Imaizumi, A.; Mori, M.; Suzuki, H.; Uo, M.; Hashimoto, M.; Naganuma, M.; Matsuoka, K.; Mizuno, S.; et al. Decreased Plasma Histidine Level Predicts Risk of Relapse in Patients with Ulcerative Colitis in Remission. PLoS One 2015, 10, e0140716. [Google Scholar] [CrossRef]
- Liu, W.; Liu, T.; Yin, M. Beneficial effects of histidine and carnosine on ethanol-induced chronic liver injury. Food Chem. Toxicol. 2008, 46, 1503–1509. [Google Scholar] [CrossRef]
- Yan, S.; Wu, S.; Yin, M.; Chen, H.; Chen, H. Protective Effects from Carnosine and Histidine on Acetaminophen-Induced Liver Injury. J. Food Sci. 2009, 74, H259–H265. [Google Scholar] [CrossRef]
- Hisatsune, T.; Kaneko, J.; Kurashige, H.; Cao, Y.; Satsu, H.; Totsuka, M.; Katakura, Y.; Imabayashi, E.; Matsuda, H. Effect of Anserine/Carnosine Supplementation on Verbal Episodic Memory in Elderly People. J. Alzheimer’s Dis. 2015, 50, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Baye, E.; Menon, K.; de Courten, M.P.; Earnest, A.; Cameron, J.; de Courten, B. Does supplementation with carnosine improve cardiometabolic health and cognitive function in patients with pre-diabetes and type 2 diabetes? study protocol for a randomised, double-blind, placebo-controlled trial. BMJ Open 2017, 7, e017691. [Google Scholar] [CrossRef]
- Sahin, S.; Burukoglu Donmez, D. Effects of Carnosine (Beta-Alanyl-L-Histidine) in an Experimental Rat Model of Acute Kidney Injury Due to Septic Shock. Med. Sci. Monit. 2018, 24, 305–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Case, J.; Khan, S.; Khalid, R.; Khan, A. Epidemiology of Acute Kidney Injury in the Intensive Care Unit. Crit. Care Res. Pract. 2013, 2013, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Wu, Q.; Zhang, X.; He, Q.; Zhao, H. Mechanistic Evaluation of the Protective Effect of Carnosine on Acute Lung Injury in Sepsis Rats. Pharmacology 2017, 100, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Furst, T.; Massaro, A.; Miller, C.; Williams, B.T.; LaMacchia, Z.M.; Horvath, P.J. β-Alanine supplementation increased physical performance and improved executive function following endurance exercise in middle aged individuals. J. Int. Soc. Sports Nutr. 2018, 15, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glenn, J.M.; Gray, M.; Stewart, R.; Moyen, N.E.; Kavouras, S.A.; DiBrezzo, R.; Turner, R.; Baum, J. Incremental effects of 28 days of beta-alanine supplementation on high-intensity cycling performance and blood lactate in masters female cyclists. Amino Acids 2015, 47, 2593–2600. [Google Scholar] [CrossRef]
- Glenn, J.M.; Gray, M.; Stewart, R.W.; Moyen, N.E.; Kavouras, S.A.; DiBrezzo, R.; Turner, R.; Baum, J.I.; Stone, M.S. Effects of 28-Day Beta-Alanine Supplementation on Isokinetic Exercise Performance and Body Composition in Female Masters Athletes. J. Strength Cond. Res. 2016, 30, 200–207. [Google Scholar] [CrossRef]
- del Favero, S.; Roschel, H.; Solis, M.Y.; Hayashi, A.P.; Artioli, G.G.; Otaduy, M.C.; Benatti, F.B.; Harris, R.C.; Wise, J.A.; Leite, C.C.; et al. Beta-alanine (CarnosynTM) supplementation in elderly subjects (60–80 years): Effects on muscle carnosine content and physical capacity. Amino Acids 2012, 43, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Small, B.J.; Rawson, K.S.; Martin, C.; Eisel, S.L.; Sanberg, C.D.; McEvoy, C.L.; Sanberg, P.R.; Shytle, R.D.; Tan, J.; Bickford, P.C. Nutraceutical Intervention Improves Older Adults’ Cognitive Functioning. Rejuvenation Res. 2014, 17, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Caruso, G.; Godos, J.; Castellano, S.; Micek, A.; Murabito, P.; Galvano, F.; Ferri, R.; Grosso, G.; Caraci, F. The Therapeutic Potential of Carnosine/Anserine Supplementation against Cognitive Decline: A Systematic Review with Meta-Analysis. Biomedicines 2021, 9, 253. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Baba, S.P.; Sweeney, B.R.; Barski, O.A. Detoxification of aldehydes by histidine-containing dipeptides: From chemistry to clinical implications. Chem. Biol. Interact. 2013, 202, 288–297. [Google Scholar] [CrossRef] [Green Version]
- Charoensin, S.; Laopaiboon, B.; Boonkum, W.; Phetcharaburanin, J.; Villareal, M.O.; Isoda, H.; Duangjinda, M. Thai Native Chicken as a Potential Functional Meat Source Rich in Anserine, Anserine/Carnosine, and Antioxidant Substances. Animals 2021, 11, 902. [Google Scholar] [CrossRef]
- Kai, S.; Watanabe, G.; Kubota, M.; Kadowaki, M.; Fujimura, S. Effect of dietary histidine on contents of carnosine and anserine in muscles of broilers. Anim. Sci. J. 2015, 86, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Tomonaga, S.; Matsumoto, M.; Furuse, M. β-Alanine Enhances Brain and Muscle Carnosine Levels in Broiler Chicks. J. Poult. Sci. 2012, 49, 308–312. [Google Scholar] [CrossRef] [Green Version]
- Kralik, G.; Kralik, Z.; Djurkin Kušec, I.; Škrtić, Z.; Kralik, I. Influence of Dietary Histidine, Hybrid Line and Gender on Chicken Meat Quality and Carnosine Concentration. J. Poult. Sci. 2015, 52, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Kralik, G.; Sak-Bosnar, M.; Kralik, Z.; Galović, O.; Grčević, M.; Kralik, I. Effect of β-alanine and L-histidine on concentration of carnosine in muscle tissue and oxidative stability of chicken meat. Poljoprivreda 2015, 21, 190–194. [Google Scholar] [CrossRef]
- Haug, A.; R⊘dbotten, R.; Mydland, L.T.; Christophersen, O.A. Increased broiler muscle carnosine and anserine following histidine supplementation of commercial broiler feed concentrate. Acta Agric. Scand. Sect. A Anim. Sci. 2008, 58, 71–77. [Google Scholar] [CrossRef]
- Harris, R.C.; Tallon, M.J.; Dunnett, M.; Boobis, L.; Coakley, J.; Kim, H.J.; Fallowfield, J.L.; Hill, C.A.; Sale, C.; Wise, J.A. The absorption of orally supplied β-alanine and its effect on muscle carnosine synthesis in human vastus lateralis. Amino Acids 2006, 30, 279–289. [Google Scholar] [CrossRef]
- Kopec, W.; Jamroz, D.; Wiliczkiewicz, A.; Biazik, E.; Pudlo, A.; Korzeniowska, M.; Hikawczuk, T.; Skiba, T. Antioxidative Characteristics of Chicken Breast Meat and Blood after Diet Supplementation with Carnosine, L-histidine, and β-alanine. Antioxidants 2020, 9, 1093. [Google Scholar] [CrossRef]
- Qi, B.; Wang, J.; Ma, Y.; Wu, S.; Qi, G.; Zhang, H. Effect of dietary β-alanine supplementation on growth performance, meat quality, carnosine content, and gene expression of carnosine-related enzymes in broilers. Poult. Sci. 2018, 97, 1220–1228. [Google Scholar] [CrossRef] [PubMed]
- Perim, P.; Marticorena, F.M.; Ribeiro, F.; Barreto, G.; Gobbi, N.; Kerksick, C.; Dolan, E.; Saunders, B. Can the Skeletal Muscle Carnosine Response to Beta-Alanine Supplementation Be Optimized? Front. Nutr. 2019, 6. [Google Scholar] [CrossRef] [Green Version]
- Cong, J.; Zhang, L.; Li, J.; Wang, S.; Gao, F.; Zhou, G. Effects of dietary supplementation with carnosine on growth performance, meat quality, antioxidant capacity and muscle fiber characteristics in broiler chickens. J. Sci. Food Agric. 2017, 97, 3733–3741. [Google Scholar] [CrossRef] [PubMed]
- Kralik, G.; Sak-Bosnar, M.; Grčević, M.; Kralik, Z. Effect of Amino Acids on Growth Performance, Carcass Characteristics, Meat Quality, and Carnosine Concentration in Broiler Chickens. J. Poult. Sci. 2018, 55, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Soyer, A.; Özalp, B.; Dalmış, Ü.; Bilgin, V. Effects of freezing temperature and duration of frozen storage on lipid and protein oxidation in chicken meat. Food Chem. 2010, 120, 1025–1030. [Google Scholar] [CrossRef]
- Cong, J.; Zhang, L.; Li, J.; Wang, S.; Gao, F.; Zhou, G. Effects of dietary supplementation with carnosine on meat quality and antioxidant capacity in broiler chickens. Br. Poult. Sci. 2017, 58, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Petersmann, A.; Müller-Wieland, D.; Müller, U.A.; Landgraf, R.; Nauck, M.; Freckmann, G.; Heinemann, L.; Schleicher, E. Definition, Classification and Diagnosis of Diabetes Mellitus. Exp. Clin. Endocrinol. Diabetes 2019, 127, S1–S7. [Google Scholar] [CrossRef] [Green Version]
- Soliman, K.M.; Mohamed, A.M.; Metwally, N.S. Attenuation of Some Metabolic Deteriorations Induced by Diabetes Mellitus Using Carnosine. J. Appl. Sci. 2007, 7, 2252–2260. [Google Scholar] [CrossRef] [Green Version]
- Kubomura, D.; Matahira, Y.; Nagai, K.; Niijima, A. Effect of anserine ingestion on hyperglycemia and the autonomic nerves in rats and humans. Nutr. Neurosci. 2010, 13, 183–188. [Google Scholar] [CrossRef]
- Sauerhofer, S.; Yuan, G.; Braun, G.S.; Deinzer, M.; Neumaier, M.; Gretz, N.; Floege, J.; Kriz, W.; van der Woude, F.; Moeller, M.J. L-Carnosine, a Substrate of Carnosinase-1, Influences Glucose Metabolism. Diabetes 2007, 56, 2425–2432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Sawalha, N.A.; Alshogran, O.Y.; Awawdeh, M.S.; Almomani, B.A. The effects of l-Carnosine on development of metabolic syndrome in rats. Life Sci. 2019, 237, 116905. [Google Scholar] [CrossRef]
- Albrecht, T.; Schilperoort, M.; Zhang, S.; Braun, J.D.; Qiu, J.; Rodriguez, A.; Pastene, D.O.; Krämer, B.K.; Köppel, H.; Baelde, H.; et al. Carnosine Attenuates the Development of both Type 2 Diabetes and Diabetic Nephropathy in BTBR ob/ob Mice. Sci. Rep. 2017, 7, 44492. [Google Scholar] [CrossRef] [PubMed]
- Peters, V.; Schmitt, C.P.; Zschocke, J.; Gross, M.-L.; Brismar, K.; Forsberg, E. Carnosine treatment largely prevents alterations of renal carnosine metabolism in diabetic mice. Amino Acids 2012, 42, 2411–2416. [Google Scholar] [CrossRef]
- Janssen, B.; Hohenadel, D.; Brinkkoetter, P.; Peters, V.; Rind, N.; Fischer, C.; Rychlik, I.; Cerna, M.; Romzova, M.; de Heer, E.; et al. Carnosine as a Protective Factor in Diabetic Nephropathy: Association With a Leucine Repeat of the Carnosinase Gene CNDP1. Diabetes 2005, 54, 2320–2327. [Google Scholar] [CrossRef] [Green Version]
- Fatih Aydın, A.; Küçükgergin, C.; Bingül, İ.; Doğan-Ekici, I.; Doğru-Abbasoğlu, S.; Uysal, M. Effect of Carnosine on Renal Function, Oxidation and Glycation Products in the Kidneys of High-Fat Diet/Streptozotocin-Induced Diabetic Rats. Exp. Clin. Endocrinol. Diabetes 2017, 125, 282–289. [Google Scholar] [CrossRef]
- Forsberg, E.A.; Botusan, I.R.; Wang, J.; Peters, V.; Ansurudeen, I.; Brismar, K.; Catrina, S.B. Carnosine decreases IGFBP1 production in db/db mice through suppression of HIF-1. J. Endocrinol. 2015, 225, 159–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karkabounas, S.; Papadopoulos, N.; Anastasiadou, C.; Gubili, C.; Peschos, D.; Daskalou, T.; Fikioris, N.; Simos, Y.V.; Kontargiris, E.; Gianakopoulos, X.; et al. Effects of α -Lipoic Acid, Carnosine, and Thiamine Supplementation in Obese Patients with Type 2 Diabetes Mellitus: A Randomized, Double-Blind Study. J. Med. Food 2018, 21, 1197–1203. [Google Scholar] [CrossRef]
- Peng, W.; Mao, P.; Liu, L.; Chen, K.; Zhong, Y.; Xia, W.; Guo, Q.; Tan, S.C.; Rahmani, J.; Kord Varkaneh, H.; et al. Effect of carnosine supplementation on lipid profile, fasting blood glucose, HbA1C and insulin resistance: A systematic review and meta-analysis of long-term randomized controlled trials. Complement. Ther. Med. 2020, 48, 102241. [Google Scholar] [CrossRef]
- Caruso, G.; Fresta, C.G.; Grasso, M.; Santangelo, R.; Lazzarino, G.; Lunte, S.M.; Caraci, F. Inflammation as the Common Biological Link Between Depression and Cardiovascular Diseases: Can Carnosine Exert a Protective Role? Curr. Med. Chem. 2020, 27, 1782–1800. [Google Scholar] [CrossRef]
- Gimbrone, M.A.; García-Cardeña, G. Endothelial Cell Dysfunction and the Pathobiology of Atherosclerosis. Circ. Res. 2016, 118, 620–636. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Li, Y.; Li, Y.; Ren, X.; Zhang, X.; Hu, D.; Gao, Y.; Xing, Y.; Shang, H. Oxidative Stress-Mediated Atherosclerosis: Mechanisms and Therapies. Front. Physiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Caruso, G.; Fresta, C.G.; Fidilio, A.; O’Donnell, F.; Musso, N.; Lazzarino, G.; Grasso, M.; Amorini, A.M.; Tascedda, F.; Bucolo, C.; et al. Carnosine Decreases PMA-Induced Oxidative Stress and Inflammation in Murine Macrophages. Antioxidants 2019, 8, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, M.-A.M.; Builta, Z.J.; Monroe, T.B.; Doorn, J.A.; Anderson, E.J. Biochemical characterization of the catecholaldehyde reactivity of l-carnosine and its therapeutic potential in human myocardium. Amino Acids 2019, 51, 97–102. [Google Scholar] [CrossRef]
- Lombardi, C.; Carubelli, V.; Lazzarini, V.; Vizzardi, E.; Bordonali, T.; Ciccarese, C.; Castrini, A.I.; Dei Cas, A.; Nodari, S.; Metra, M. Effects of oral administration of orodispersible levo-carnosine on quality of life and exercise performance in patients with chronic heart failure. Nutrition 2015, 31, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Kumral, A.; Giriş, M.; Soluk-Tekkeşin, M.; Olgaç, V.; Doğru-Abbasoğlu, S.; Türkoğlu, Ü.; Uysal, M. Beneficial effects of carnosine and carnosine plus vitamin E treatments on doxorubicin-induced oxidative stress and cardiac, hepatic, and renal toxicity in rats. Hum. Exp. Toxicol. 2016, 35, 635–643. [Google Scholar] [CrossRef]
- Gan, L.; Johnson, J.A. Oxidative damage and the Nrf2-ARE pathway in neurodegenerative diseases. Biochim. Biophys. Acta Mol. Basis Dis. 2014, 1842, 1208–1218. [Google Scholar] [CrossRef] [Green Version]
- Bhat, A.H.; Dar, K.B.; Anees, S.; Zargar, M.A.; Masood, A.; Sofi, M.A.; Ganie, S.A. Oxidative stress, mitochondrial dysfunction and neurodegenerative diseases; a mechanistic insight. Biomed. Pharmacother. 2015, 74, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Cahill-Smith, S.; Li, J.-M. Oxidative stress, redox signalling and endothelial dysfunction in ageing-related neurodegenerative diseases: A role of NADPH oxidase 2. Br. J. Clin. Pharmacol. 2014, 78, 441–453. [Google Scholar] [CrossRef] [Green Version]
- Cojocaru, I.M.; Cojocaru, M.; Sapira, V.; Ionescu, A. Evaluation of oxidative stress in patients with acute ischemic stroke. Rom. J. Intern. Med. 2013, 51, 97–106. [Google Scholar]
- Fendri, C.; Mechri, A.; Khiari, G.; Othman, A.; Kerkeni, A.; Gaha, L. Implication du stress oxydant dans la physiopathologie de la schizophrénie: Revue de la literature. Encephale. 2006, 32, 244–252. [Google Scholar] [CrossRef]
- Maria Michel, T.; Pulschen, D.; Thome, J. The Role of Oxidative Stress in Depressive Disorders. Curr. Pharm. Des. 2012, 18, 5890–5899. [Google Scholar] [CrossRef] [Green Version]
- Fedorova, T.N.; Belyaev, M.S.; Trunova, O.A.; Gnezditsky, V.V.; Maximova, M.Y.; Boldyrev, A.A. Neuropeptide carnosine increases stability of lipoproteins and red blood cells as well as efficiency of immune competent system in patients with chronic discirculatory encephalopathy. Biochem. Suppl. Ser. A Membr. Cell Biol. 2009, 3, 62–65. [Google Scholar] [CrossRef]
- Boldyrev, A.; Fedorova, T.; Stepanova, M.; Dobrotvorskaya, I.; Kozlova, E.; Boldanova, N.; Bagyeva, G.; Ivanova-Smolenskaya, I.; Illarioshkin, S. Carnisone Increases Efficiency of DOPA Therapy of Parkinson’s Disease: A Pilot Study. Rejuvenation Res. 2008, 11, 821–827. [Google Scholar] [CrossRef]
- Davis, C.K.; Laud, P.J.; Bahor, Z.; Rajanikant, G.; Majid, A. Systematic review and stratified meta-analysis of the efficacy of carnosine in animal models of ischemic stroke. J. Cereb. Blood Flow Metab. 2016, 36, 1686–1694. [Google Scholar] [CrossRef] [Green Version]
- Yasuhara, T.; Hara, K.; Maki, M.; Masuda, T.; Sanberg, C.D.; Sanberg, P.R.; Bickford, P.C.; Borlongan, C.V. Dietary Supplementation Exerts Neuroprotective Effects in Ischemic Stroke Model. Rejuvenation Res. 2008, 11, 201–214. [Google Scholar] [CrossRef]
- Wang, A.; Ma, Q.; Wang, X.; Xu, G. Protective effects of beef decoction rich in carnosine on cerebral ischemia injury by permanent middle cerebral artery occlusion in rats. Exp. Ther. Med. 2017. [Google Scholar] [CrossRef]
- Pekcetin, C.; Kiray, M.; Ergur, B.; Tugyan, K.; Bagriyanik, H.; Erbil, G.; Baykara, B.; Camsari, U. Carnosine attenuates oxidative stress and apoptosis in transient cerebral ischemia in rats. Acta Biol. Hung. 2009, 60, 137–148. [Google Scholar] [CrossRef]
- Wang, J.-P.; Yang, Z.-T.; Liu, C.; He, Y.-H.; Zhao, S.-S. L-carnosine inhibits neuronal cell apoptosis through signal transducer and activator of transcription 3 signaling pathway after acute focal cerebral ischemia. Brain Res. 2013, 1507, 125–133. [Google Scholar] [CrossRef]
- Min, J.; Senut, M.-C.; Rajanikant, K.; Greenberg, E.; Bandagi, R.; Zemke, D.; Mousa, A.; Kassab, M.; Farooq, M.U.; Gupta, R.; et al. Differential neuroprotective effects of carnosine, anserine, and N -acetyl carnosine against permanent focal ischemia. J. Neurosci. Res. 2008, 86, 2984–2991. [Google Scholar] [CrossRef] [Green Version]
- Dichter, M.A. Emerging Insights Into Mechanisms of Epilepsy: Implications for New Antiepileptic Drug Development. Epilepsia 1994, 35, S51–S57. [Google Scholar] [CrossRef] [PubMed]
- Gietzen, D.W.; Lindström, S.H.; Sharp, J.W.; Teh, P.S.; Donovan, M.J. Indispensable Amino Acid–Deficient Diets Induce Seizures in Ketogenic Diet–Fed Rodents, Demonstrating a Role for Amino Acid Balance in Dietary Treatments for Epilepsy. J. Nutr. 2018, 148, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Qi, Z.; Yu, X.; Xu, P.; Hao, Y.; Pan, X.; Zhang, C. L-Homocarnosine, l-carnosine, and anserine attenuate brain oxidative damage in a pentylenetetrazole-induced epilepsy model of ovariectomized rats. 3 Biotech 2018, 8, 363. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Ding, M.; Zhu-Ge, Z.-B.; Zhu, Y.-Y.; Jin, C.; Chen, Z. Carnosine, a precursor of histidine, ameliorates pentylenetetrazole-induced kindled seizures in rat. Neurosci. Lett. 2006, 400, 146–149. [Google Scholar] [CrossRef]
- Zhu, Y.-Y.; Zhu-Ge, Z.-B.; Wu, D.-C.; Wang, S.; Liu, L.-Y.; Ohtsu, H.; Chen, Z. Carnosine inhibits pentylenetetrazol-induced seizures by histaminergic mechanisms in histidine decarboxylase knock-out mice. Neurosci. Lett. 2007, 416, 211–216. [Google Scholar] [CrossRef]
- Kozan, R.; Sefil, F.; Bağırıcı, F. Anticonvulsant effect of carnosine on penicillin-induced epileptiform activity in rats. Brain Res. 2008, 1239, 249–255. [Google Scholar] [CrossRef]
- Jin, H.-B.; Li, B.; Cu, J.; Cheng, J.-S.; Yang, R. Electro-acupuncture improves epileptic seizures induced by kainic acid in taurine-depletion rats. Acupunct. Electrother. Res. 2005, 30, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Kamiński, R.M.; Żółkowska, D.; Kozicka, M.; Kleinrok, Z.; Czuczwar, S.J. L-Histidine is a beneficial adjuvant for antiepileptic drugs against maximal electroshock-induced seizures in mice. Amino Acids 2004, 26, 85–89. [Google Scholar] [CrossRef]
- Feng, Z.; Zheng, X.; Wang, J. Effects of carnosine on the evoked potentials in hippocampal CA1 region. J. Zhejiang Univ. Sci. B 2009, 10, 505–511. [Google Scholar] [CrossRef] [Green Version]
- Guiotto, A.; Calderan, A.; Ruzza, P.; Borin, G. Carnosine and Carnosine-Related Antioxidants: A Review. Curr. Med. Chem. 2005, 12, 2293–2315. [Google Scholar] [CrossRef]
- Koffie, R.M.; Hyman, B.T.; Spires-Jones, T.L. Alzheimer’s disease: Synapses gone cold. Mol. Neurodegener. 2011, 6, 63. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Green, B.D. Temporal Effects of Neuron-specific beta-secretase 1 (BACE1) Knock-in on the Mouse Brain Metabolome: Implications for Alzheimer’s Disease. Neuroscience 2019, 397, 138–146. [Google Scholar] [CrossRef] [Green Version]
- Sase, A.; Dahanayaka, S.; Höger, H.; Wu, G.; Lubec, G. Changes of hippocampal beta-alanine and citrulline levels are paralleling early and late phase of retrieval in the Morris Water Maze. Behav. Brain Res. 2013, 249, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, J.; Enya, A.; Enomoto, K.; Ding, Q.; Hisatsune, T. Anserine (beta-alanyl-3-methyl-L-histidine) improves neurovascular-unit dysfunction and spatial memory in aged AβPPswe/PSEN1dE9 Alzheimer’s-model mice. Sci. Rep. 2017, 7, 12571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corona, C.; Frazzini, V.; Silvestri, E.; Lattanzio, R.; La Sorda, R.; Piantelli, M.; Canzoniero, L.M.T.; Ciavardelli, D.; Rizzarelli, E.; Sensi, S.L. Effects of Dietary Supplementation of Carnosine on Mitochondrial Dysfunction, Amyloid Pathology, and Cognitive Deficits in 3xTg-AD Mice. PLoS ONE 2011, 6, e17971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Busquets, X.; Ponce, J.; Bravo, R.; Arimon, M.; Martianez, T.; Gella, A.; Cladera, J.; Durany, N. Modulation of amyloid β peptide1-42 cytotoxicity and aggregation in vitro by glucose and chondroitin sulfate. Curr. Alzheimer Res. 2010, 7, 428–438. [Google Scholar] [CrossRef]
- Loske, C.; Gerdemann, A.; Schepl, W.; Wycislo, M.; Schinzel, R.; Palm, D.; Riederer, P.; Münch, G. Transition metal-mediated glycoxidation accelerates cross-linking of β-amyloid peptide. Eur. J. Biochem. 2000, 267, 4171–4178. [Google Scholar] [CrossRef]
- Colín-Barenque, L.; Bizarro-Nevares, P.; González Villalva, A.; Pedraza-Chaverri, J.; Medina-Campos, O.N.; Jimenez-Martínez, R.; Rodríguez-Rangel, D.S.; Reséndiz, S.; Fortoul, T.I. Neuroprotective effect of carnosine in the olfactory bulb after vanadium inhalation in a mouse model. Int. J. Exp. Pathol. 2018, 99, 180–188. [Google Scholar] [CrossRef]
- Kenneth Maiese; Zhao Zhong Chong; Yan Chen Shang Mechanistic Insights Into Diabetes Mellitus and Oxidative Stress. Curr. Med. Chem. 2007, 14, 1729–1738. [CrossRef]
- Ma, J.; Chen, J.; Bo, S.; Lu, X.; Zhang, J. Protective effect of carnosine after chronic cerebral hypoperfusion possibly through suppressing astrocyte activation. Am. J. Transl. Res. 2015, 7, 2706–2715. [Google Scholar]
- Hata, J.; Ohara, T.; Katakura, Y.; Shimizu, K.; Yamashita, S.; Yoshida, D.; Honda, T.; Hirakawa, Y.; Shibata, M.; Sakata, S.; et al. Association Between Serum β-Alanine and Risk of Dementia. Am. J. Epidemiol. 2019, 188, 1637–1645. [Google Scholar] [CrossRef]
- Masuoka, N.; Yoshimine, C.; Hori, M.; Tanaka, M.; Asada, T.; Abe, K.; Hisatsune, T. Effects of Anserine/Carnosine Supplementation on Mild Cognitive Impairment with APOE4. Nutrients 2019, 11, 1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuoka, N.; Lei, C.; Li, H.; Inamura, N.; Shiotani, S.; Yanai, N.; Sato, K.; Sakurai, K.; Hisatsune, T. Anserine, HClO-scavenger, protected against cognitive decline in individuals with mild cognitive impairment. Aging 2021, 13, 1729–1741. [Google Scholar] [CrossRef]
- Cornelli, U. Treatment of Alzheimer’s Disease with a Cholinesterase Inhibitor Combined with Antioxidants. Neurodegener. Dis. 2010, 7, 193–202. [Google Scholar] [CrossRef]
- Rybakova, Y.S.; Boldyrev, A.A. Effect of Carnosine and Related Compounds on Proliferation of Cultured Rat Pheochromocytoma PC-12 Cells. Bull. Exp. Biol. Med. 2012, 154, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Oppermann, H.; Schnabel, L.; Meixensberger, J.; Gaunitz, F. Pyruvate attenuates the anti-neoplastic effect of carnosine independently from oxidative phosphorylation. Oncotarget 2016, 7, 85848–85860. [Google Scholar] [CrossRef] [Green Version]
- Renner, C.; Asperger, A.; Seyffarth, A.; Meixensberger, J.; Gebhardt, R.; Gaunitz, F. Carnosine inhibits ATP production in cells from malignant glioma. Neurol. Res. 2010, 32, 101–105. [Google Scholar] [CrossRef]
- Hipkiss, A.R.; Gaunitz, F. Inhibition of tumour cell growth by carnosine: Some possible mechanisms. Amino Acids 2014, 46, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Oppermann, H.; Faust, H.; Yamanishi, U.; Meixensberger, J.; Gaunitz, F. Carnosine inhibits glioblastoma growth independent from PI3K/Akt/mTOR signaling. PLoS ONE 2019, 14, e0218972. [Google Scholar] [CrossRef] [Green Version]
- Hwang, B.; Shin, S.-S.; Song, J.-H.; Choi, Y.H.; Kim, W.-J.; Moon, S.-K. Carnosine exerts antitumor activity against bladder cancers in vitro and in vivo via suppression of angiogenesis. J. Nutr. Biochem. 2019, 74, 108230. [Google Scholar] [CrossRef]
- WU, C.-C.; LAI, P.-Y.; HSIEH, S.; CHENG, C.-C.; HSIEH, S.-L. Suppression of Carnosine on Adhesion and Extravasation of Human Colorectal Cancer Cells. Anticancer Res. 2019, 39, 6135–6144. [Google Scholar] [CrossRef]
- Iovine, B.; Oliviero, G.; Garofalo, M.; Orefice, M.; Nocella, F.; Borbone, N.; Piccialli, V.; Centore, R.; Mazzone, M.; Piccialli, G.; et al. The Anti-Proliferative Effect of L-Carnosine Correlates with a Decreased Expression of Hypoxia Inducible Factor 1 alpha in Human Colon Cancer Cells. PLoS ONE 2014, 9, e96755. [Google Scholar] [CrossRef] [Green Version]
- Iovine, B.; Guardia, F.; Irace, C.; Bevilacqua, M.A. L-carnosine dipeptide overcomes acquired resistance to 5-fluorouracil in HT29 human colon cancer cells via downregulation of HIF1-alpha and induction of apoptosis. Biochimie 2016, 127, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Mikuła-Pietrasik, J.; Książek, K. L-Carnosine Prevents the Pro-cancerogenic Activity of Senescent Peritoneal Mesothelium Towards Ovarian Cancer Cells. Anticancer Res. 2016, 36, 665–671. [Google Scholar]
- Ignarro, L.J.; Napoli, C. Novel features of nitric oxide, endothelial nitric oxide synthase, and atherosclerosis. Curr. Diab. Rep. 2005, 5, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Alaghband-Zadeh, J.; Mehdizadeh, S.; Khan, N.S.; Farrell, A.O.; Bitensky, L.; Chayen, J. The natural substrate for nitric oxide synthase activity. Cell Biochem. Funct. 2001, 19, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Nakashima, Y.; Toda, K. Carnosine Facilitates Nitric Oxide Production in Endothelial F-2 Cells. Biol. Pharm. Bull. 2009, 32, 1836–1839. [Google Scholar] [CrossRef] [Green Version]
- Severina, I.S.; Bussygina, O.G.; Pyatakova, N.V. Carnosine as a regulator of soluble guanylate cyclase. Biochemistry 2000, 65, 783–788. [Google Scholar]
- Nicoletti, V.G.; Santoro, A.M.; Grasso, G.; Vagliasindi, L.I.; Giuffrida, M.L.; Cuppari, C.; Purrello, V.S.; Stella, A.M.G.; Rizzarelli, E. Carnosine interaction with nitric oxide and astroglial cell protection. J. Neurosci. Res. 2007, 85, 2239–2245. [Google Scholar] [CrossRef]
- O’Dowd, A.; O’Dowd, J.J.; Miller, D.J. The dipeptide carnosine constricts rabbit saphenous vein as a zinc complex apparently via a serotonergic receptor. J. Physiol. 1996, 495, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Ririe, D.G.; Roberts, P.R.; Shouse, M.N.; Zaloga, G.P. Vasodilatory actions of the dietary peptide carnosine. Nutrition 2000, 16, 168–172. [Google Scholar] [CrossRef]
- Vanhoutte, P.M. Serotonin and the vascular wall. Int. J. Cardiol. 1987, 14, 189–203. [Google Scholar] [CrossRef]
- Nagai, K.; Tanida, M.; Niijima, A.; Tsuruoka, N.; Kiso, Y.; Horii, Y.; Shen, J.; Okumura, N. Role of l-carnosine in the control of blood glucose, blood pressure, thermogenesis, and lipolysis by autonomic nerves in rats: Involvement of the circadian clock and histamine. Amino Acids 2012, 43, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Mong, M.; Chao, C.; Yin, M. Histidine and carnosine alleviated hepatic steatosis in mice consumed high saturated fat diet. Eur. J. Pharmacol. 2011, 653, 82–88. [Google Scholar] [CrossRef]
- Liu, Y.; Cotillard, A.; Vatier, C.; Bastard, J.-P.; Fellahi, S.; Stévant, M.; Allatif, O.; Langlois, C.; Bieuvelet, S.; Brochot, A.; et al. A Dietary Supplement Containing Cinnamon, Chromium and Carnosine Decreases Fasting Plasma Glucose and Increases Lean Mass in Overweight or Obese Pre-Diabetic Subjects: A Randomized, Placebo-Controlled Trial. PLoS ONE 2015, 10, e0138646. [Google Scholar] [CrossRef] [PubMed]
- Versari, D.; Daghini, E.; Virdis, A.; Ghiadoni, L.; Taddei, S. Endothelial Dysfunction as a Target for Prevention of Cardiovascular Disease. Diabetes Care 2009, 32, S314–S321. [Google Scholar] [CrossRef] [Green Version]
- Ihara, H.; Kakihana, Y.; Yamakage, A.; Kai, K.; Shibata, T.; Nishida, M.; Yamada, K.; Uchida, K. 2-Oxo-histidine–containing dipeptides are functional oxidation products. J. Biol. Chem. 2019, 294, 1279–1289. [Google Scholar] [CrossRef] [Green Version]
- Prokopieva, V.D.; Yarygina, E.G.; Bokhan, N.A.; Ivanova, S.A. Use of Carnosine for Oxidative Stress Reduction in Different Pathologies. Oxid. Med. Cell. Longev. 2016, 2016, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Daiber, A.; Steven, S.; Weber, A.; Shuvaev, V.V.; Muzykantov, V.R.; Laher, I.; Li, H.; Lamas, S.; Münzel, T. Targeting vascular (endothelial) dysfunction. Br. J. Pharmacol. 2017, 174, 1591–1619. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Deproteinization | Extraction Method | Separation Media | pH | Ref. |
|---|---|---|---|---|---|
| muscle | perchloric acid | SPE | borate buffer | 9.6 | [32,33] |
| muscle | n.a. | Liquid. | borate buffer | 9.6 | [34] |
| muscle | n.a. | Liquid | water | n.a. | [35] |
| muscle | 70% ethanol | Liquid. | water/NaOH, HCl | n.a. | [36] |
| muscle | 35% sulfosalicylic acid | Liquid. | borate buffer | n.a. | [37,38] |
| muscle | acetonitrile | Liquid. | hydrochloric acid | n.a. | [39] |
| human plasma | 35% sulfosalicylic acid | Liquid. | borate buffer | n.a. | [28,37] |
| human plasma | 35% sulfosalicylic acid | Liquid. | PBS buffer | n.a. | [18] |
| human plasma | methanol | Liquid | water/formic acid | n.a. | [40] |
| human plasma/urine | EDTA | Liquid | sodium acetate buffer | 6.4 | [41] |
| Protocol | Column | Detection | Derivatization Reagent | Sample Matrix | Limits of Detection | Ref. |
|---|---|---|---|---|---|---|
| RP-HPLC | Hypersil ODS | Fluorescent | 3-mercaptopropionic acid/o-phthaldehyde | muscle | 0.005 mmol/kg | [32,33] |
| RP-HPLC | APEX ODS | Fluorescent | 3-mercaptopropionic acid/o-phthaldehyde | muscle | 0.005 mmol/kg | [34] |
| RP HPLC | Hypersil ODS | Fluorescent | 3-mercaptopropionic acid/o-phthaldehyde | muscle | 0.005 mmol/kg | [35] |
| JLC-300 | LCR-6 | n.a | n.a. | muscle | 0.005 mmol/kg | [36] |
| RP HPLC | XBridge BEH | Fluorescent | AccQTag | muscle | n.a. | [38] |
| RP HPLC | Hypersilica | UV (210 nm) | n.a. | muscle | 3 and 10 μM | [41] |
| RP HPLC | HILIC silica | UV (214 nm) | n.a | muscle | [39] | |
| RP HPLC | Hypersil ODS | Fluorescent | 3-mercaptopropionic acid/o-phthaldehyde | human plasma | [18] | |
| UPLC | Acquity Peptide BEH C18 | MS | n.a. | human plasma | [40] | |
| RP HPLC | Jupiter C18 | Fluorescent | carbazole-9-carbonyl chloride (CFC) | human serum, animal feed | 15 nM | [28] |
| Treatments | Content of Carnosine | ||
|---|---|---|---|
| Breast | Thigh | ||
| Kralik et.al. (2015) [160] | β-alanine 0.0% | 756.15 | - |
| L-histidine 0.0% | 941.58 | - | |
| β-alanine 0.5% | 753.29 | - | |
| L-histidine 0.3% | 1025.22 | - | |
| β-alanine 1.0% | 911.01 | - | |
| L-histidine 0.5% | 1186.06 | - | |
| Kralik et. al. (2018) [168] | Control | 665.47 | 261.19 |
| β-alanine 0.5% + MgO 0.24% | 715.45 | 420.64 | |
| L-histidine 0.25% + MgO 0.24% | 736.17 | 467.40 | |
| β-alanine 0.20% + L-histidine 0.10% + MgO 0.24% | 1084.25 | 495.01 | |
| Study Design | Daily Intake of Carnosine | Main Findings of Carnosine Effects |
|---|---|---|
| Rats (n = 40) received conventional diet (control), high-fat high-carbohydrate diet, carnosine and conventional diet, or carnosine and high-fat high-carbohydrate diet [175] | 250 mg/kg/daily intraperitoneal; 16 weeks | -reduced abdominal obesity, blood pressure, glucose; -normalized total cholesterol, LDL level -no effect on insulin, leptin and adiponectin concentrations |
| BTBR (Black and Tan, BRachyuric) ob/ob mice (n = 35), a type 2 diabetes model with a phenotype like advanced human diabetes nephropathy [176] | 45 mg/kg body weight dissolved in drinking water | -improved glucose metabolism, albuminuria and restored the glomerular ultrastructure |
| Effect of carnosine on renal function, oxidation and glycation products in the kidneys of high-fat diet/streptozotocin-induced diabetic rats (n = 24) [179] | 250 mg/kg body weight; intraperitoneal, 5 times a week; 4 weeks | -decreased serum lipids, creatinine, and urea levels, oxidation products of lipids and proteins, advanced glycation end products (AGEs) levels |
| Study Design | Daily Intake of Carnosine | Main Findings of Carnosine Effects |
|---|---|---|
| Obese type 2 diabetic patients (n = 82) were either supplemented daily with α-lipoic acid, carnosine and thiamine [181] | 7 mg α-lipoic acid/kg body weight, 6 mg carnosine/kg body weight, and 1 mg thiamine/kg body weight or placebo for 8 weeks | -reduced glucose and HbA1c levels, probably by increasing insulin production from pancreas |
| Patients with type 2 diabetes mellitus (n = 54) divided into two groups, received either placebo or carnosine [129] | L carnosine 2 capsules of 500 mg each for 12 weeks. | -decrease in fat mass, fasting blood glucose, glycated hemoglobin and serum levels of triglycerides |
| Study Design | Daily Intake of Carnosine/Cells Exposure with Carnosine | Main Findings of Carnosine Effects |
|---|---|---|
| Carnosine was tested for its ability to counteract oxidative stress in macrophages [186] | Carnosine (5, 10, 20 mM) | -multimodal mechanism of action on macrophage cells under oxidative stress and inflammation conditions |
| Mitochondria from myocardial atrial samples were isolated and incubated for 3 h at 37 °C with 75-µM norepinephrine NE) and increasing concentrations of carnosine (1, 2.5, 5, 10, and 25 mM) [187] | Carnosine (1 mM) | -carnosine block formation of catecholaldehyde protein adducts in isolated human cardiac mitochondria treated with NE |
| Rats (n = 24) were treated with carnosine or carnosine + vitamin E. On the 8th day of treatment, rats were injected with a single dose of doxorubicin [189] | Carnosine 250 mg/kg/day i.p. or carnosine + vit E (200 mg/kg) once every 3 days; i.m.); for 12 days | -carnosine and especially in combination with vitamin E, protect against doxorubicin-induced toxicity in heart, liver, and kidney tissues of rats |
| Patients (n = 50) with stable chronic heart failure (CHF) and severe left-ventricular systolic dysfunction on optimal medical therapy were randomized 1:1 to receive oral or dispersible carnosine or standard treatment [188] | Carnosine 500 mg once a day; 6 months | -beneficial effects on exercise performance and quality of life in stable CHF |
| Study Design | Tumors | Cells Exposure with Carnosine | Main Findings of Carnosine Effects |
|---|---|---|---|
| Experimental Two cell lines (U87 and T98 G) [232] | Glioblastoma | Carnosine (50 mM), the PI3K inhibitor Ly-294,002 (5 μM), the mTORC1 inhibitor rapamycin (25 nM) and combinations of the compounds for 24 h | -reduces Akt phosphorylation in the U87 cell line |
| Experimental In EJ bladder cancer cells and EJ-xenografted BALB/c nude mice (n = 7 per each group) [233] | Bladder cancer | Carnosine (0,10, 20, 50, and 100 mM) for 24 h. Carnosine (0, 5, 10 mg/kg) were administered by oral gavage daily. Then the efficacy was compared with that of 5 mg/kg cisplatin, the positive control. | -stops the G1 phase cell cycle, suppresses the binding of transcription factors on the MMP-9 promoter -in xenograft tumors exhibited antitumor activity equivalent to cisplatin, but no weight loss occurred in carnosine-treated mice. |
| Experimental The HCT-116 human colorectal cancer cell line The EA.hy926 human umbilical vein endothelial cell line Cells treated with sterile water served as the control [235,236] | Colorectal cancer | Carnosine (0.5, 1 or 5 mM) was added EA.hy926 cells for 12, 24, 48 h. HCT-116 cells were treated with 1 μg/mL lipopolysaccharide, then 0.5, 1 or 5 mM carnosine combined with 1 μg/mL LPS was added and cells were incubated for 24 h | -suppress adhesion of HCT-116 cells to EA.hy926 cells and extravasation. -inhibits the NF-kB signaling pathway activation -reduced the permeability of EA.hy926 cell–cell junctions -inhibited HCT-116 cell adhesion to EA.hy926 cells |
| Experimental Primary human peritoneal mesothelial cells, three ovarian cancer cell lines: A2780, OVCAR-3 and SKOV-3 [237] | Ovarian cancer | L-carnosine (20 mM) | -inhibits mitochondria-related oxidative stress |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jukić, I.; Kolobarić, N.; Stupin, A.; Matić, A.; Kozina, N.; Mihaljević, Z.; Mihalj, M.; Šušnjara, P.; Stupin, M.; Ćurić, Ž.B.; et al. Carnosine, Small but Mighty—Prospect of Use as Functional Ingredient for Functional Food Formulation. Antioxidants 2021, 10, 1037. https://doi.org/10.3390/antiox10071037
Jukić I, Kolobarić N, Stupin A, Matić A, Kozina N, Mihaljević Z, Mihalj M, Šušnjara P, Stupin M, Ćurić ŽB, et al. Carnosine, Small but Mighty—Prospect of Use as Functional Ingredient for Functional Food Formulation. Antioxidants. 2021; 10(7):1037. https://doi.org/10.3390/antiox10071037
Chicago/Turabian StyleJukić, Ivana, Nikolina Kolobarić, Ana Stupin, Anita Matić, Nataša Kozina, Zrinka Mihaljević, Martina Mihalj, Petar Šušnjara, Marko Stupin, Željka Breškić Ćurić, and et al. 2021. "Carnosine, Small but Mighty—Prospect of Use as Functional Ingredient for Functional Food Formulation" Antioxidants 10, no. 7: 1037. https://doi.org/10.3390/antiox10071037
APA StyleJukić, I., Kolobarić, N., Stupin, A., Matić, A., Kozina, N., Mihaljević, Z., Mihalj, M., Šušnjara, P., Stupin, M., Ćurić, Ž. B., Selthofer-Relatić, K., Kibel, A., Lukinac, A., Kolar, L., Kralik, G., Kralik, Z., Széchenyi, A., Jozanović, M., Galović, O., ... Drenjančević, I. (2021). Carnosine, Small but Mighty—Prospect of Use as Functional Ingredient for Functional Food Formulation. Antioxidants, 10(7), 1037. https://doi.org/10.3390/antiox10071037